

V体育ios版 - Abstract
AMP-activated protein kinase (AMPK) is an important regulator of cellular energy status. In adipocytes, stimuli that increase intracellular cyclic AMP (cAMP) have also been shown to increase the activity of AMPK. The precise molecular mechanisms responsible for cAMP-induced AMPK activation are not clear VSports最新版本. Phosphodiesterase 3B (PDE3B) is a critical regulator of cAMP signalling in adipocytes. Here we investigated the roles of PDE3B, PDE4, protein kinase B (PKB) and the exchange protein activated by cAMP 1 (Epac1), as well as lipolysis, in the regulation of AMPK in primary rat adipocytes. We demonstrate that the increase in phosphorylation of AMPK at T172 induced by the adrenergic agonist isoproterenol can be diminished by co-incubation with insulin. The diminishing effect of insulin on AMPK activation was reversed upon treatment with the PDE3B specific inhibitor OPC3911 but not with the PDE4 inhibitor Rolipram. Adenovirus-mediated overexpression of PDE3B and constitutively active PKB both resulted in greatly reduced isoproterenol-induced phosphorylation of AMPK at T172. Co-incubation of adipocytes with isoproterenol and the PKA inhibitor H89 resulted in a total ablation of lipolysis and a reduction in AMPK phosphorylation/activation. Stimulation of adipocytes with the Epac1 agonist 8-pCPT-2’O-Me-cAMP led to increased phosphorylation of AMPK at T172. The general lipase inhibitor Orlistat decreased isoproterenol-induced phosphorylation of AMPK at T172. This decrease corresponded to a reduction of lipolysis from adipocytes. Taken together, these data suggest that PDE3B and PDE4 regulate cAMP pools that affect the activation/phosphorylation state of AMPK and that the effects of cyclic AMP on AMPK involve Epac1, PKA and lipolysis.
Keywords: PDE3B, PDE4, Epac, AMPK, PKA, lipolysis, cAMP, adipocytes
1. Introduction
AMP-activated protein kinase (AMPK) is a critical regulator of energy homeostasis. It functions as a cellular energy sensor, responding to increases in the AMP:ATP ratio by inactivating energy consuming processes such as lipogenesis and cholesterol biosynthesis, while activating energy producing processes such as fatty acid oxidation [1]. AMPK is a heterotrimeric enzyme with alpha, beta and gamma subunits, the alpha subunit being the catalytic subunit [2]. Activation of AMPK is known to be mediated by phosphorylation of the catalytic subunit at threonine 172 (T172) by the upstream kinases LKB1 and Ca2+-calmodulin kinase kinase β [3-5]. It is also thought that regulation of AMPK activity is increased directly by the binding of AMP to the regulatory subunit which also makes the holoenzyme resistant to dephosphorylation by phosphatases [6]. It has been shown that ischemia, oxidative stress and exercise increase the cellular AMP:ATP ratio and activate AMPK [7-9]. It has also been demonstrated in cultured adipocytes that agents which increase intracellular cAMP also increase the activity of AMPK [10, 11]. In contrast, in cultured pancreatic β-cells agents that stimulate cAMP production resulted in decreased AMPK activity [12]. Although many of the downstream effects of cAMP are mediated by cAMP dependant protein kinase (PKA), exchange proteins directly activated by cAMP (Epac1 and Epac2) have more recently been shown to mediate signals downstream of cAMP independent of PKA [13, 14] VSports注册入口. Also, in some cell types, cAMP and cGMP mediate effects by binding to cyclic nucleotide gated ion channels [15]. The intracellular level of cAMP is regulated by the balance between production of cAMP by adenylate cyclase and the breakdown of cAMP by the actions of the 11 member superfamily of cyclic nucleotide phosphodiesterases (PDE1-11). Phosphodiesterase enzymes differ in their primary structure, affinities for cAMP and cGMP, responses to specific effectors and inhibitors, and mechanisms through which they are regulated [16].
PDE3B and PDE4 enzymes are the most abundant PDEs in adipocytes. Whereas little is known regarding the role of PDE4s in adipocyte metabolic functions, the role and regulation of PDE3B have been extensively studied [17, 18]. Results from knockout and transgenic mouse models, as well as results from isolated primary adipocytes and β-cells demonstrate an important role of PDE3B in the regulation of glucose and lipid metabolism, in insulin secretion and in adipocyte functions [19] [20, 21] V体育官网入口. For example, in adipocytes, insulin-induced phosphorylation and activation of PDE3B, mediated via protein kinase B (PKB) [22, 23], is the major mechanism whereby insulin inhibits catecholamine-induced lipolysis. Furthermore, PDE3B plays an important role in the regulation of insulin-induced glucose uptake and lipogenesis in adipocytes [21].
It has been recently shown that lipolysis is at least partially required for the cAMP-induced AMPK activation in 3T3-L1 adipocytes [24]. The re-esterification of fatty acids that occurs during lipolysis leads to a depletion of ATP and an increase in the AMP:ATP ratio, thus activating AMPK [24]. Further studies have demonstrated that fatty acids themselves can lead to an increased activation of AMPK in HepG2 cells [25]. The mechanisms involved in the regulation of AMPK in response to changes in the intracellular level of cAMP in primary adipocytes are at this point unclear. It is currently believed that cAMP exists in cells as discrete pools, each with local expression of proteins that regulate, and are regulated by, cAMP VSports在线直播. In this study we investigate the role of PDE3B, PDE4, PKA, PKB, Epac and lipolysis in the phosphorylation and activation of AMPK in primary adipocytes.
2. Materials and Methods
2.1 Animals
Male Sprague-Dawley rats (from B&K Universal Stockholm, Sweden) between 36 and 42 weeks of age were used for the isolation of primary adipocytes. All experiments have been approved by the Animal Ethics Committee, Lund, Sweden V体育2025版.
2.2 Chemicals and Reagents
Isoproterenol (SIGMA), Insulin (Novo Nordisk) and the Epac agonist 8-pCPT-2’-O-Me-cAMP (Biolog life-science institute) were used for the treatment of adipocytes. The PDE3 inhibitor OPC3911 (Otsuka pharmaceuticals), general lipase inhibitor Orlistat (kind gift of Dr VSports. Maria Sörhede Winzell), PKA inhibitor H89 and PDE4 inhibitor Rolipram (both from Biomol international) were used as inhibitors. All antibodies used for immunoblotting, with the exception of anti-PDE3B (raised in-house in rabbits against the peptide CGYYGSGKMFRRPSLP from PDE3B) and total PKB (a kind gift from Dario Alessi, University of Dundee, raised against a peptide corresponding to residues 466-480 of rat PKBα), were from Cell Signaling Technologies. AMPKα1 antibody used for the AMPK activity assays was a kind gift of Professor Grahame Hardie, University of Dundee (U. K. ). The peptide AMARAASAAALRRR (AMARA) used for the AMPK activity assays was synthesized by Dr. Graham Bloomberg, University of Bristol (U. K. ).
V体育官网 - 2.3 Isolation of primary adipocytes
Primary rat adipocytes were isolated from epididymal fat pads as previously described [26]. Adipocytes were diluted to a concentration of 10% (v/v) in Krebs Ringer (KRH) buffer containing 25 mM Hepes, 200 nM adenosine, 2 mM glucose and 1% bovine serum albumin (BSA). One milliliter of cell suspension was incubated for 37 °C as indicated in Results. Incubations were stopped by washing the adipocytes in BSA free KRH buffer and homogenization in a buffer containing 50 mM TES, 2 mM EGTA, 1 mM EDTA, 250 mM sucrose, 40 mM phenylphosphate, 5 mM NaF, 1 mM dithioerythriol, 0. 5 mM sodium orthovanadate, 10 μg/mL antipain, 10 μg/mL leupeptin, 1 μg/mL pepstatin A, pH 7. 4 VSports app下载. Homogenates were centrifuged at 5000 × g for 5 minutes at 4 °C, the fat cake removed and the supernatants solublized in 1% (v/v) Nonidet P40 on ice for 15 minutes. Solublized lysates were centrifuged at 10000 × g for 10 minutes at 4 °C and protein concentrations were determined by the method of Bradford [27].
2.4 Adenovirus mediated overexpression of PDE3B and constitutively active PKB
Primary rat adipocytes (2.5 mL of 12.5% (v/v)) were infected with high titer virus stocks (1×1010 PFU/mL) encoding β-Galactosidase (Ad-β-GAL), FLAG-PDE3B (Ad-PDE3B), or myristoylated PKB (Ad-PKBmyr) adenoviral vectors. Infection of cells was carried out in DMEM containing 5% fetal calf serum, 0.5% BSA, 100 U/mL penicillin and 100 μg/mL streptomycin at 37 °C overnight with 5% CO2. After infection cells were washed and suspended in KRH buffer. Aliquots of 800 μL of cell suspension were incubated as described in results for 30 minutes at 37 °C before washing and homogenized as described above. Homogenates were centrifuged at 5000 × g for 5 minutes at 4 °C, the fat cake removed and the supernatants solublized with a 1/10 volume of 10% (v/v) Nonidet P40 on ice for 15 minutes. Solublized lysates were centrifuged at 10000 × g for 10 minutes at 4 °C and the supernatants stored at -20 °C. Protein concentrations were determined by the method of Bradford [27]. The Ad-β-GAL adenoviral vector was kindly provided by Drs. C.N. Newgard and H. Mulder. Ad-PKBmyr was a kind gift of Dr. Michael Quon. The Ad-PDE3B adenoviral vector was produced in house.
V体育安卓版 - 2.5 Lipolysis
Primary rat adipocytes were incubated as described above (Sec 2.3). After incubation for specified times, cell suspensions were centrifuged at 500 × g for 30 seconds, and the supernatants collected for measurement of glycerol content, as described by Dole [28]. Briefly, buffer containing 50 mM glycine, 0.05% hydrazine hydrate, 1 mM MgCl2, 0.75 mg/mL ATP, 0.375 mg/mL NAD, 25 μg/mL glycerol-3-phosphate dehydrogenase and 0.5 μg/mL glycerokinase was added to the cell-free supernatants. The reaction was incubated at room temperature for 40 minutes and OD340 was measured. Glycerol release from adipocytes was calculated as μmol glycerol released per ten minutes per mL of reaction volume.
2.6 SDS-PAGE and Western Blotting
Samples were mixed with Laemmli sample buffer, subjected to electrophoresis through 7% polyacrylamide gels, and subsequently transferred to Hybond C nitrocellulose membranes (Amersham biosciences). Membranes were blocked with 10% milk in TBST (50 mM Tris pH 7.5, 150 mM NaCl and 0.1% (w/v) Tween-20) followed by incubation with primary antibody, as indicated in results, overnight at 4 °C. Membranes were washed and subsequently incubated for 1 hour at room temperature with a 1:10,000 dilution of HRP conjugated-anti-rabbit IgG secondary antibody (Biosource) in 5% milk. After a second washing, membranes were incubated with ECL reagent (Pierce) for 5 minutes before image capture in a LAS-1000 (Fujifilm) CCD camera. Western blots were quantified using Image Guage software (Fujifilm).
VSports - 2.7 AMPK activity assay
Cell lysates (10 μg of protein) were incubated at 4°C for 1h on a shaking platform with 1 μg of anti-AMPKα1 antibody conjugated to 5 μl of protein G-Sepharose (GE Healthcare Biosciences). The immunoprecipitates were washed twice with 0.5 mL of Lysis Buffer (50 mM Tris-HCL pH 7.5, 1 mM EGTA, 1 mM EDTA, 1 mM Na-orthovanadate, 50 mM NaF, 5 mM pyrophosphate, 270 mM Sucrose, 1% (w/v) Nonidet P40, 0.5 M NaCl and 1 mM DTT) and twice with 0.5 ml of 50 mM Tris-HCl pH 7.5, 0.1 mM EGTA and 1 mM DTT (Buffer A). Phosphotransferase activity towards the AMARA peptide was then measured in a total assay volume of 50 μl containing 50 mM Tris-HCl pH 7.5, 0.1% (v/v) 2-mercaptoethanol, 10 mM MgCl2, 0.1 mM EGTA, 0.1 mM [γ-32P]-ATP (300 cpm/pmol) and 200 μM AMARA peptide for 20 min at 30°C. Incorporation of 32P-phosphate into the peptide substrate was determined by applying 40 μl of the reaction mixture onto P81 phosphocellulose paper, followed by washing of the papers in 50 mM phosphoric acid and scintillation counting. One Unit (U) of activity was defined as that which catalyzed the incorporation of 1 nmol of 32P/min into the substrate.
VSports手机版 - 2.8 Data analysis
Data are expressed as the mean values ± SEM of the indicated number of experiments. Statistical significance (here as p < 0.05) was determined using the unpaired Student’s t test and shown as stars in the figures.
"VSports最新版本" 3. Results
"V体育平台登录" 3.1 PDE3B and PDE4 are involved in the regulation of AMPK
Activation of AMPK has previously been demonstrated to correlate with phosphorylation of T172 in the activation loop of the α subunit [29]. Using an antibody specific for AMPKα phosphorylated at T172, we were able to determine the activation state of AMPK under different experimental conditions. To determine whether PDE3B is involved in cAMP and insulin-mediated regulation of AMPK, adipocytes were treated with different combinations of insulin, isoproterenol, a β-adrenergic receptor agonist which stimulates the production of intracellular cAMP, and OPC3911, a specific inhibitor for PDE3. As can be seen in Figure 1a, stimulation of primary rat adipocytes with isoproterenol resulted in a four-fold increase in AMPK phosphorylation. This isoproterenol-induced AMPK phosphorylation was reduced by 56 % upon co-treatment with insulin. Treatment of adipocytes with a combination of insulin, isoproterenol and the PDE3 inhibitor OPC3911 resulted in a reversal of the effect of insulin on isoproterenol-induced AMPK phosphorylation. The effect of insulin on isoproterenol-induced AMPK phosphorylation was not reversed after pretreatment with the PDE4 inhibitor Rolipram. The phosphorylation of acetyl CoA carboxylase (ACC), a downstream target of AMPK, at S79 closely matched that of AMPK in each of the experimental conditions. Thus, PDE3B is the major phosphodiesterase mediating the inhibitory effect of insulin on isoproterenol-induced AMPK phosphorylation in adipocytes. As can also be seen in Figure 1a, treatment of adipocytes with inhibitors of PDE3 and PDE4 each resulted in an increased basal AMPK phosphorylation, suggesting a role for both PDEs in the regulation of AMPK in the absence of hormonal stimulation. When measuring the phosphorylation state of proteins that contain PKA phosphorylation consensus sequences it is clear that both PDE3 and PDE4 inhibitors increase basal phosphorylation of PKA targets as well as potentiate isoproterenol induced phosphorylation of PKA target proteins (Figure 1b).
Fig. 1.
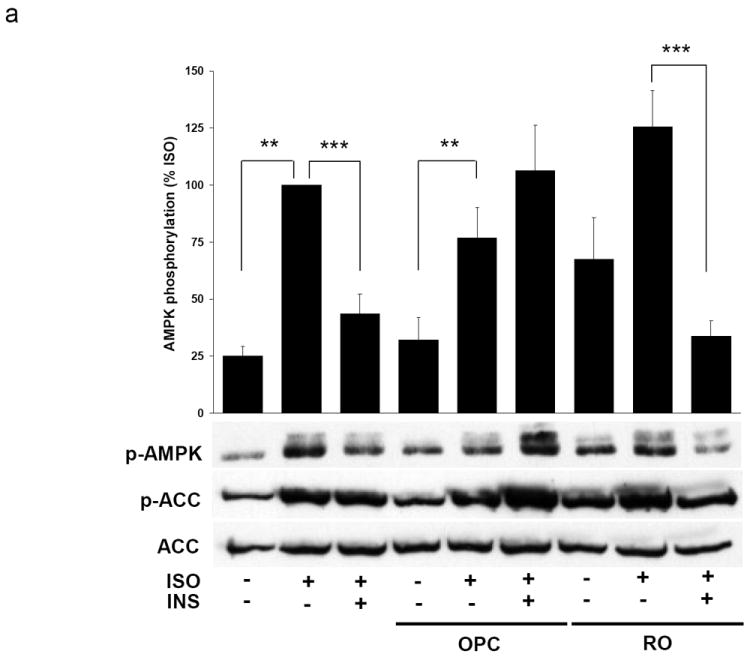

PDE3B and PDE4 are involved in the regulation of AMPK Primary adipocytes were stimulated with 30 nM isoproterenol (ISO) alone or in combination with 1nM insulin (INS) for 10 minutes. Adipocytes were pre-treated for 10 minutes with OPC3911 (OPC) or Rolipram (RO) prior to stimulation with hormones as indicated. Representative immunoblots for A) phospho-AMPK T172, phospho-ACC S79 and total ACC and B) for phospho-PKA substrates are shown. Data is presented as the mean ± SEM of the quantified phosphorylation relative to the isoproterenol stimulated sample. n = 3-7. ** p<0.01, *** p<0.001.
3.2 Overexpression of PDE3B and constitutively active PKB results in reduced isoproterenol induced AMPK activation (VSports手机版)
Insulin-induced phosphorylation and activation of PDE3B has been shown to be mediated via the upstream kinase PKB [22, 23, 30]. In order to obtain further evidence regarding a role of PDE3B in mediating insulin-induced lowering of cAMP mediated phosphorylation of AMPK we overexpressed constitutively active PKB (Ad-PKBmyr) and wild type PDE3B (Ad-PDE3B), respectively, using an adenovirus system, and studied their effects on AMPK phosphorylation. As shown in Figure 2a, adipocytes overexpressing PDE3B showed a reduced AMPK phosphorylation in response to isoproterenol at all concentrations tested, as compared to control adipocytes (adipocytes overexpressing β-Galactosidase). Adipocytes overexpressing PDE3B displayed less than 50% of the isoproterenol-induced AMPK phosphorylation, when stimulated with 30 μM or 100 μM isoproterenol, as compared to control cells. Similar to overexpressed PDE3B, overexpression of constitutively active PKB resulted in a reduction of isoproterenol-induced AMPK phosphorylation, when adipocytes were treated with 30 μM and 100 μM isoproterenol (Figure 2b). The reductions in AMPK phosphorylation were reflected in the phosphorylation of ACC, which was also reduced in adipocytes overexpressing both PDE3B and constitutively active PKB, in response to isoproterenol.
Fig. 2.


Overexpression of PDE3B and constitutively active PKB results in reduced isoproterenol-induced AMPK activation Primary adipocytes were infected overnight with adenoviruses containing β-Galactosidase (Ad-β-Gal) and A) FLAG-PDE3B (Ad-PDE3B) or B) myristoylated PKB (Ad-PKBmyr) adenoviral vectors. Infected adipocytes were stimulated with 3, 10, 30 or 100 nM isoproterenol (ISO) for 30 minutes prior to homogenization. Aliquots of homogenates were used for western blotting. Representative western blots for phosphor-AMPK T172, phosphor-ACC S79, total ACC and PDE3B or total PKB are shown. Data is presented as mean ± SEM relative to the unstimulated sample in each group. n = 3-4. * p<0.05, *** p<0.001.
3.3 PKA and Epac1 mediate isoproterenol- induced activation of AMPK
We next investigated whether protein kinase A (PKA), which has an important role in the phosphorylation and activation of hormone sensitive lipase (HSL), and thereby activation of lipolysis, was required for the activation of AMPK by cAMP. Co-treatment of primary adipocytes with isoproterenol and the PKA inhibitor H89 resulted in inhibition of lipolysis as well as AMPK phosphorylation/activation in a dose-dependent manner, however, the dose-dependencies for these effects were different (Figure 3). Thus, 25 μM H89 inhibited lipolysis by 71% (Figure 3c) with no inhibition of AMPK activity (Figure 3b). At higher concentrations, H89 completely blocked both lipolysis and phosphorylation of HSL at the regulatory site S563, whereas AMPK phosphorylation/activation was inhibited to a much lesser extent (Figure 3a and c). We next wanted to determine whether the effect of cAMP on AMPK activation could be partly mediated through the PKA-independent Epac1 signaling pathway. It has been shown that the Epac1 agonist 8-pCPT-2’-O-Me-cAMP elicits a decrease in insulin-induced lipogenesis and an increase in phosphorylation of acetyl-CoA carboxylase, a downstream target of AMPK, in primary rat adipocytes [31]. Stimulation of primary adipocytes with the Epac1 agonist 8-pCPT-2’-O-Me-cAMP resulted in increases in AMPK phosphorylation and activity (Figure 4a and b) with minimal activation of lipolysis or phosphorylation of HSL as determined by measurement of glycerol release from adipocytes (Figure 4c).
Fig. 3.


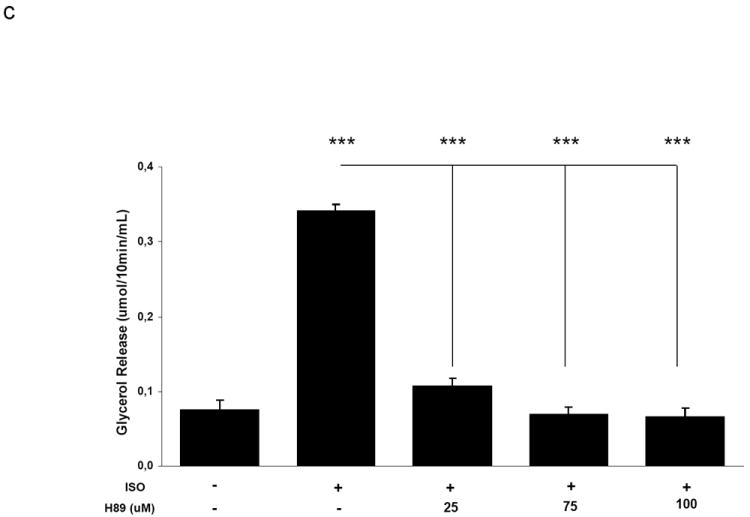
Activation of AMPK by isoproterenol is partially mediated by PKA. Adipocytes were pre-treated with vehicle (DMSO) or H89 at the indicated concentrations for 30 minutes followed by stimulation with 30nM isoproterenol for 10 minutes. Aliquots of homogenates were used for A) western blotting and B) AMPK activity assays. C) Prior to homogenizations, the incubation media was collected for measurement of glycerol content. Data is presented as the mean ± SEM relative to the isoproterenol stimulated sample unless indicated. n = 4. * p<0.05, *** p<0.001.
Fig. 4.



Activation of Epac1 results in increased AMPK phosphorylation and activity. Adipocytes were incubated in the absence or presence of 10 μM of the Epac agonist 8-pCPT-2’-O-Me-cAMP (Epac Ag) for 40 minutes. As a control untreated adipocytes were incubated for 30 minutes followed by 10 minutes of stimulation with 30 nM isoproterenol. Prior to homogenizations, the incubation media were collected for measurement of glycerol content. Data is presented as the mean ± SEM. n = 7. * p<0.05, *** p<0.001.
3.4 The general lipase inhibitor Orlistat reduces isoproterenol-mediated AMPK phosphorylation in primary rat adipocytes
It has been demonstrated in 3T3-L1 adipocytes that the ATP-utilizing fatty acid re-esterification process that occurs during lipolysis results in subsequent alteration of the intracellular AMP:ATP ratio and is at least partially responsible for isoproterenol-induced increase in AMPK activity [24]. As shown in Figure 5a, inhibition of lipolysis with Orlistat blunted the effect of isoproterenol on AMPK phosphorylation in primary rat adipocytes. While Orlistat treatment alone resulted in a slight increase in AMPK phosphorylation, it reduced the isoproterenol effect on AMPK phosphorylation in adipocytes stimulated with 30 and 100 nM isoproterenol. The reduction in AMPK phosphorylation was also reflected in the phosphorylation of ACC at S79, as well as HSL at S563. This corresponded with a 37 % decrease in isoproterenol-induced glycerol release from adipocytes in all isoproterenol stimulated adipocytes pre-treated with Orlistat (Figure 5b).
Fig. 5.


The general lipase inhibitor Orlistat reduces isoproterenol-mediated AMPK activation in primary rat adipocytes. A) Primary adipocytes were pre-treated with either vehicle (60% DMSO, 40% ethanol) or 100 μM Orlistat for 30 minutes followed by stimulation with 0, 10, 30 or 100 nM isoproterenol for 10 minutes. Aliquots of homogenates were used for western blotting. B) Prior to homogenizations, the incubation media was collected for measurement of glycerol content. Data is presented as mean ± SEM relative to the unstimulated sample. n = 4. * p<0.05, ** p<0.01.
V体育官网 - 4. Discussion
For the first time, in this study we demonstrate that PDE3B and its upstream regulator PKB, as well as PDE4, are involved in the regulation of cAMP pools of importance for AMPK phosphorylation. Furthermore, downstream of cAMP, several important regulators of AMPK phosphorylation were identified; PKA, Epac1 and lipolysis.
Whereas PDE3B as well as PDE4 appear to control pools of cAMP regulating AMPK phosphorylation in the absence of insulin, PDE3B, but not PDE4, mediates insulin-induced lowering of AMPK phosphorylation. The lack of effect of PDE4 in this context is not due to an inefficient inhibition of PDE4 by Rolipram since Rolipram treatment per se resulted in increased basal AMPK phosphorylation and a potentiation of isoproterenol-induced AMPK phosphorylation. Furthermore, using PKA consensus sequence antibodies, it was demonstrated that Rolipram pre-treatment resulted in increased basal as well as a potentiation of isoproterenol-mediated phosphorylation of adipocyte proteins. In agreement with the finding that PDE3 inhibition increases basal and blocks insulin-induced phosphorylation of AMPK, overexpression of PDE3B and a constitutively active form of PKB resulted in reduction of isoproterenol-induced AMPK phosphorylation. Another PKB-dependent mechanism responsible for mediating insulin-induced lowering of AMPK phosphorylation at T172 involves PKB-induced phosphorylation of AMPK at Serine 485, which corresponds to a decrease in AMPK activity in perfused rat hearts [32]. Whether this mode of regulation exists in adipocytes remains to be established. However, the complete reversal of AMPK phosphorylation upon treatment with PDE3 inhibitor suggests a major role for PDE3B in mediating insulin-induced inhibition of cAMP-dependent AMPK phosphorylation. cAMP initiates the activation of multiple signalling pathways including, but not limited to, the PKA and Epac signalling pathways. The finding that isoproterenol-induced AMPK phosphorylation/activation and lipolysis showed different dose response curves after treatment with H89, with AMPK phosphorylation being less sensitive to inhibition with regard to dose and maximal effects as compared to lipolysis, indicates the involvement of additional, PKA-independent, mechanisms for AMPK phosphorylation. Thus, we investigated whether Epac signalling could be involved in regulating AMPK phosphorylation in adipocytes. Epac1 was previously demonstrated to be expressed in rat adipocytes and was shown to affect insulin-induced lipogenesis and phosphorylation of ACC [31]. Using the Epac1 agonist 8-pCPT-2’-Me-cAMP we showed PKA-independent phosphorylation of AMPK suggesting that PKA as well as Epac1 contribute to isoproterenol-induced phosphorylation of AMPK. Epac proteins are known to activate the Rap family of small GTPases [14]. It has recently been demonstrated that the Epac1-Rap1 axis is critical for adipocyte differentiation and other important signalling pathways in adipocytes [33]. The finding of increased phosphorylation of AMPK in response to Epac activation in our study provides further evidence of an important role for Epac in mediation of the effects of cAMP on adipocyte homeostasis.
The finding that pre-treatment of primary adipocytes with the general lipase inhibitor Orlistat inhibited isoproterenol-induced AMPK activation is in agreement with previous findings in 3T3-L1 adipocytes [24]. It was suggested that re-esterification of fatty acids that occurs during lipolysis depletes ATP and increases the AMP:ATP ratio, thus activating AMPK [24]. Further studies have demonstrated that fatty acids themselves can lead to an increased activation of AMPK [25, 34]. The mechanisms whereby Orlistat on its own leads to a significant increase in AMPK phosphorylation compared to vehicle alone as noted in this study are not known. The effect of Orlistat on basal AMPK phosphorylation/activation could be related to the fact that Orlistat is a general lipase inhibitor and affects not only HSL but also adipose triglyceride lipase, diacylglyceride lipase and lipoprotein lipase. Inhibition of these lipases might have secondary effects which could lead to the activation of AMPK.
5. Conclusions
We have demonstrated that significant cross-talk occurs between the insulin, cAMP and AMPK signalling pathways in adipocytes. PKB/PDE3B appears to be at a junction of the three pathways, mediating the antagonistic effect of insulin on cAMP-induced AMPK phosphorylation. Furthermore, PDE4, PKA, Epac and lipolysis were shown to participate in controlling and mediating cAMP-induced AMPK phosphorylation/activation, respectively. Exactly what is the physiological role for this intricate crosstalk between insulin, cAMP and AMPK signalling networks needs to be further investigated.
V体育ios版 - Acknowledgments
The authors would like to acknowledge Dr. Lena Stenson for advice on the manuscript and Eva Ohlson for excellent technical assistance. Bilal Omar is a recipient of a postdoctoral fellowship from the Tage Blucher foundation. This work was supported by the Swedish Research Council project 3362 and by the Lund University Diabetes Center. Grants were obtained from the following foundations: Swedish Diabetes Association; Novo Nordisk, Denmark, the Swedish Society of Medicine, Dr. P. Håkansson and Albert Påhlsson.
Abbreviations
- PDE
phosphodiesterase
- cAMP
cyclic AMP
- Epac
exchange proteins directly activated by cAMP
- AMPK
AMP-activated protein kinase
- PKA
protein kinase A
- PKB
protein kinase B
- ACC
acetyl CoA carboxylase
- ISO
isoproterenol
- INS
insulin
V体育安卓版 - Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Carling D, Zammit VA, Hardie DG. FEBS Lett. 1987;223(2):217–222. doi: 10.1016/0014-5793(87)80292-2. [DOI] [PubMed] [Google Scholar]
- 2.Davies SP, Hawley SA, Woods A, Carling D, Haystead TA, Hardie DG. Eur J Biochem. 1994;223(2):351–357. doi: 10.1111/j.1432-1033.1994.tb19001.x. [DOI] [PubMed] [Google Scholar]
- 3.Hawley SA, Boudeau J, Reid JL, Mustard KJ, Udd L, Makela TP, Alessi DR, Hardie DG. J Biol. 2003;2(4):28. doi: 10.1186/1475-4924-2-28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Hawley SA, Pan DA, Mustard KJ, Ross L, Bain J, Edelman AM, Frenguelli BG, Hardie DG. Cell Metab. 2005;2(1):9–19. doi: 10.1016/j.cmet.2005.05.009. [DOI] [PubMed] [Google Scholar]
- 5.Woods A, Dickerson K, Heath R, Hong SP, Momcilovic M, Johnstone SR, Carlson M, Carling D. Cell Metab. 2005;2(1):21–33. doi: 10.1016/j.cmet.2005.06.005. [DOI (VSports注册入口)] [PubMed] [Google Scholar]
- 6.Sanders MJ, Ali ZS, Hegarty BD, Heath R, Snowden MA, Carling D. J Biol Chem. 2007;282(45):32539–32548. doi: 10.1074/jbc.M706543200. [DOI] [PubMed] [Google Scholar]
- 7.Beauloye C, Marsin AS, Bertrand L, Krause U, Hardie DG, Vanoverschelde JL, Hue L. FEBS Lett. 2001;505(3):348–352. doi: 10.1016/s0014-5793(01)02788-0. [VSports在线直播 - DOI] [PubMed] [Google Scholar]
- 8.Yang J, Holman GD. J Biol Chem. 2005;280(6):4070–4078. doi: 10.1074/jbc.M410213200. [DOI] [PubMed] [Google Scholar]
- 9.Winder WW, Hardie DG. Am J Physiol. 1996;270(2 Pt 1):E299–304. doi: 10.1152/ajpendo.1996.270.2.E299. [VSports最新版本 - DOI] [PubMed] [Google Scholar]
- 10.Yin W, Mu J, Birnbaum MJ. J Biol Chem. 2003;278(44):43074–43080. doi: 10.1074/jbc.M308484200. [DOI] [PubMed] [Google Scholar]
- 11.Hutchinson DS, Chernogubova E, Dallner OS, Cannon B, Bengtsson T. Diabetologia. 2005;48(11):2386–2395. doi: 10.1007/s00125-005-1936-7. [VSports app下载 - DOI] [PubMed] [Google Scholar]
- 12.Hurley RL, Barre LK, Wood SD, Anderson KA, Kemp BE, Means AR, Witters LA. J Biol Chem. 2006;281(48):36662–36672. doi: 10.1074/jbc.M606676200. [DOI (V体育安卓版)] [PubMed] [Google Scholar]
- 13.McKnight GS, Cummings DE, Amieux PS, Sikorski MA, Brandon EP, Planas JV, Motamed K, Idzerda RL. Recent Prog Horm Res. 1998;53:139–159. discussion 160-131. [PubMed] [Google Scholar]
- 14.de Rooij J, Zwartkruis FJ, Verheijen MH, Cool RH, Nijman SM, Wittinghofer A, Bos JL. Nature. 1998;396(6710):474–477. doi: 10.1038/24884. [DOI] [PubMed] [Google Scholar]
- 15.Bradley J, Reisert J, Frings S. Curr Opin Neurobiol. 2005;15(3):343–349. doi: 10.1016/j.conb.2005.05.014. [DOI] [PubMed] [Google Scholar]
- 16.Conti M, Beavo J. Annu Rev Biochem. 2007;76:481–511. doi: 10.1146/annurev.biochem.76.060305.150444. [DOI] [PubMed] [Google Scholar]
- 17.Thompson PE, Manganiello V, Degerman E. Curr Top Med Chem. 2007;7(4):421–436. doi: 10.2174/156802607779941224. [DOI] [PubMed] [Google Scholar]
- 18.Degerman E, M V, Beavo J, Francis S, Houslay MD, editors. Cyclic Nucleotide Phosphodiesterases in Health and Disease. Boca Raton, FL USA: CRC Press; 2007. pp. 79–99. [Google Scholar]
- 19.Choi YH, Park S, Hockman S, Zmuda-Trzebiatowska E, Svennelid F, Haluzik M, Gavrilova O, Ahmad F, Pepin L, Napolitano M, Taira M, Sundler F, Stenson Holst L, Degerman E, Manganiello VC. J Clin Invest. 2006;116(12):3240–3251. doi: 10.1172/JCI24867. ["VSports在线直播" DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Harndahl L, Wierup N, Enerback S, Mulder H, Manganiello VC, Sundler F, Degerman E, Ahren B, Holst LS. J Biol Chem. 2004;279(15):15214–15222. doi: 10.1074/jbc.M308952200. [DOI] [PubMed] [Google Scholar]
- 21.Zmuda-Trzebiatowska E, Oknianska A, Manganiello V, Degerman E. Cell Signal. 2006;18(3):382–390. doi: 10.1016/j.cellsig.2005.05.007. [DOI (VSports最新版本)] [PubMed] [Google Scholar]
- 22.Wijkander J, Landstrom TR, Manganiello V, Belfrage P, Degerman E. Endocrinology. 1998;139(1):219–227. doi: 10.1210/endo.139.1.5693. [DOI] [PubMed] [Google Scholar]
- 23.Kitamura T, Kitamura Y, Kuroda S, Hino Y, Ando M, Kotani K, Konishi H, Matsuzaki H, Kikkawa U, Ogawa W, Kasuga M. Mol Cell Biol. 1999;19(9):6286–6296. doi: 10.1128/mcb.19.9.6286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Gauthier MS, Miyoshi H, Souza SC, Cacicedo JM, Saha AK, Greenberg AS, Ruderman NB. J Biol Chem. 2008;283(24):16514–16524. doi: 10.1074/jbc.M708177200. [DOI (VSports注册入口)] [PMC free article] [PubMed] [Google Scholar]
- 25.Za’tara G, Bar-Tana J, Kalderon B, Suter M, Morad E, Samovski D, Neumann D, Hertz R. Biochem Pharmacol. 2008;76(10):1263–1275. doi: 10.1016/j.bcp.2008.08.028. ["VSports手机版" DOI] [PubMed] [Google Scholar]
- 26.Honnor RC, Dhillon GS, Londos C. J Biol Chem. 1985;260(28):15122–15129. [PubMed] [Google Scholar]
- 27.Bradford MM. Anal Biochem. 1976;72:248–254. doi: 10.1016/0003-2697(76)90527-3. [DOI] [PubMed] [Google Scholar]
- 28.Dole VP, Meinertz H. J Biol Chem. 1960;235:2595–2599. [PubMed] [Google Scholar]
- 29.Hawley SA, Davison M, Woods A, Davies SP, Beri RK, Carling D, Hardie DG. J Biol Chem. 1996;271(44):27879–27887. doi: 10.1074/jbc.271.44.27879. [DOI] [PubMed] [Google Scholar]
- 30.Ahmad F, Cong LN, Stenson Holst L, Wang LM, Rahn Landstrom T, Pierce JH, Quon MJ, Degerman E, Manganiello VC. J Immunol. 2000;164(9):4678–4688. doi: 10.4049/jimmunol.164.9.4678. [DOI (V体育官网入口)] [PubMed] [Google Scholar]
- 31.Zmuda-Trzebiatowska E, Manganiello V, Degerman E. Cell Signal. 2007;19(1):81–86. doi: 10.1016/j.cellsig.2006.05.024. [DOI] [PubMed] [Google Scholar]
- 32.Horman S, Vertommen D, Heath R, Neumann D, Mouton V, Woods A, Schlattner U, Wallimann T, Carling D, Hue L, Rider MH. J Biol Chem. 2006;281(9):5335–5340. doi: 10.1074/jbc.M506850200. [DOI] [PubMed] [Google Scholar]
- 33.Petersen RK, Madsen L, Pedersen LM, Hallenborg P, Hagland H, Viste K, Doskeland SO, Kristiansen K. Mol Cell Biol. 2008;28(11):3804–3816. doi: 10.1128/MCB.00709-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Watt MJ, Steinberg GR, Chen ZP, Kemp BE, Febbraio MA. J Physiol. 2006;574(Pt 1):139–147. doi: 10.1113/jphysiol.2006.107318. [DOI] [PMC free article] [PubMed] [Google Scholar]